For at least one decade I have thought about some nuclear morphology, typically found in bone marrow in progenitor cells (mouse) particularly in mice who have been dosed with 125mg/kg/day benzo(a)pyrene (BaP) for 18 days, underpinned by the presence or lack of Cyp1a1 (Fig. 3. Histology of bone marrow) which showed severe hypolasia of the bone marrow and a significant increase in the hypersegmentation of the bone marrow cells (all kinds). Lymphocytes were increased regardless of BaP in the Cyp1a1(-/-) mice regardless of BaP treatment as was hypersegmentation. So this was interesting, but never pursued as a separate topic. I have micrographs which show this same bilateral segmentation, which may turn out to be portions of (or whole) chromosome territories. I searched the literature (the relatively current literature and current literature during the study and didn’t find anyone who had any images which would suggest the cause.
Then as chromosome territories became a target of interest, two very old (I mean really old) documents were cited in Cold spring Harbor Perspectives in Biology in 2010: (Cold Spring Harb Perspect Biol. 2010 Mar;2(3):a003889. doi: 10.1101/cshperspect.a003889. Chromosome territories. Cremer T, Cremer M.) listed as Rabl 1885, and Boveri 1909 (In the drawings of two cells in prophase from Ascaris megalocephala univalens embryos. He mentioned that these protrusions were germ line chromosomes. This renewed my interest in trying to figure out which chromosomes were blipped out (or cued for hypersegmentation) in the null mice, and whether this indirectly relates simply to high levels of cell renewal in their hypodcellular bone marrow or whether it relates directly to the loss of CyP1a1 in mice somehow.
I have redrawn the figure of Boveri (1909) and added some images from bone marrow of hypersegmented cells (which I now assume to be specific chromosome territories of specific chromosome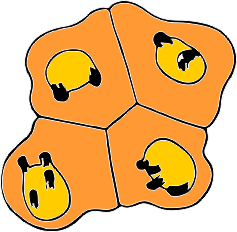 s, but which chromosomes is not known. Another interesting point is that Boveri thought that the two top cells were initially responsible for the two bottom cells (which he called daughter cells) and the difference in two vs four areas of chromosome blipping are evident (this would be high condensation and sort of a promise later of becoming an area tethered by a thin strand of nuclear membrane, or budded off as a microbody). Certainly this man spent a lot of time at the microscope. These types of symmetries, and associations are randomly seen usually and lots and lots of view-time are required before patterns emerge.
s, but which chromosomes is not known. Another interesting point is that Boveri thought that the two top cells were initially responsible for the two bottom cells (which he called daughter cells) and the difference in two vs four areas of chromosome blipping are evident (this would be high condensation and sort of a promise later of becoming an area tethered by a thin strand of nuclear membrane, or budded off as a microbody). Certainly this man spent a lot of time at the microscope. These types of symmetries, and associations are randomly seen usually and lots and lots of view-time are required before patterns emerge.
(Adding one more coincidence? comment! is that Boveri drew his cells with about a 64 degrees of “arc” from approximate (obviously not know for sure –i dont even know if anyone has decided where the “center” of the nucleus actually is.. ha ha–i will have to google that). When i calculated the angular dimensions of the two similarly painted chromosomes in FISH and other fluorescent tags for chromosome territories, i could almost always envision and measure a distance of about 70 degrees between chromosomes. This is kind of exciting, as it give bilateral symmetry to the nucleus which is NOT an exact mirror, but something different. The Arc of Separation between chromosomes I will have to research.