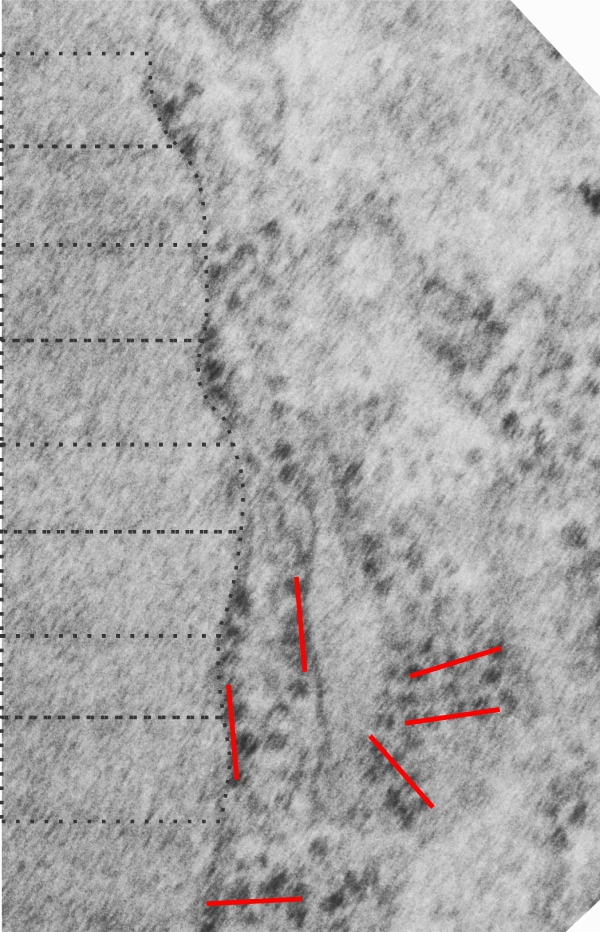
There are several samples of the banding periods and growing end ribosomes, along with a tangential portion showing a grid arrangement of ribosomes on what I presume is an RER profile that contains an abundance of surfactant protein A. The relative sizes marked in black dotted lines are the intervals seen in each separate period of the bands (visible either as 3, 5, 7, or sometimes 9 separate bands) and a bar marker in red (which is the approximate equivalent of the total period (100 nm) and a sample from the internet of a shadow cast molecule of surfactant protein A — in a tiny circle (red circle, white background) which was estimated by the authors to be about 20 nm, Relative to the width of the micrographs which I have used in this image, the single surfactant molecule is sized accordingly. Because the surfactant A protein was shadow cast it appears more defined that would be seen in reality.