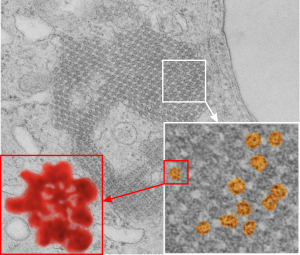
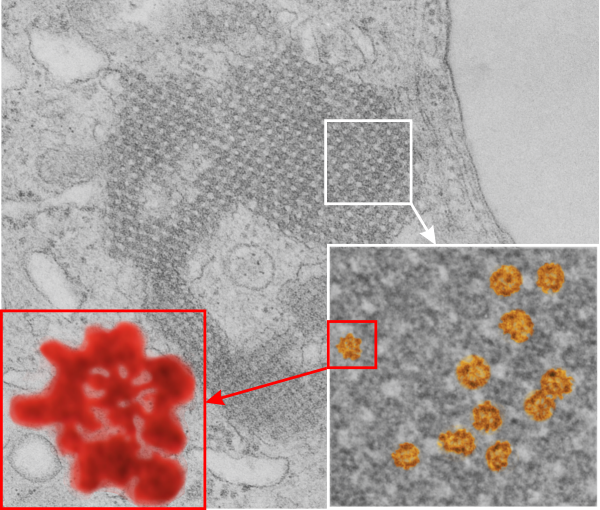
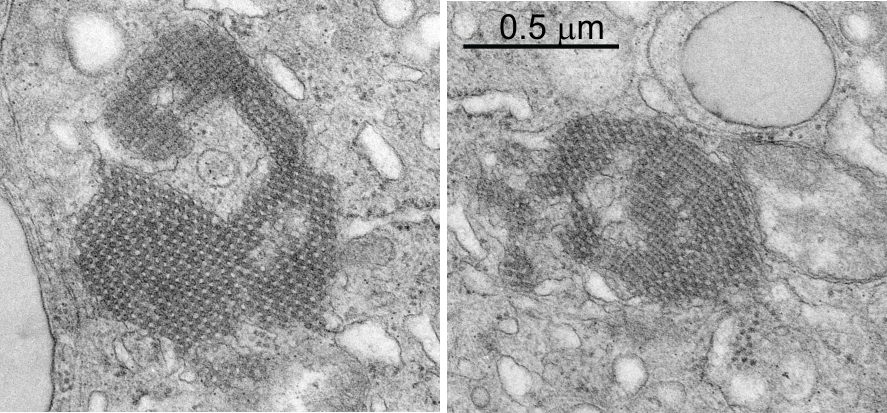
From the previous post, I did some more searching on the very ordered protein inclusion in the type Ii cell of a mongrel dog (5 yr old). The orderly array looked to me like a bunch of hexagonal structures with a central dot. When I made measurements (comparing the magnification, enlargement and pole piece used at the microscope, I figured out what about 500 nm would be, and then 50 nm seemed to be almost exactly twice as big as each of these hexagonal flower (bouquet) structures. They might not actually be octadecamers (as is often found for SP-A )- and a shadowed image of one such shadowed SP-A molecule from a publication by T. Voss, H. Eistetter, KP Schafer in J. Mol. Biol, 1988, 201: 219-227, which i have cut out, pseudocolored in photoshop, cut out, and placed along side one of the bouquets in the ordered protein inclusion in this particular cell. Since I could not identify the cell specifically, one only can guess as to what it might be.
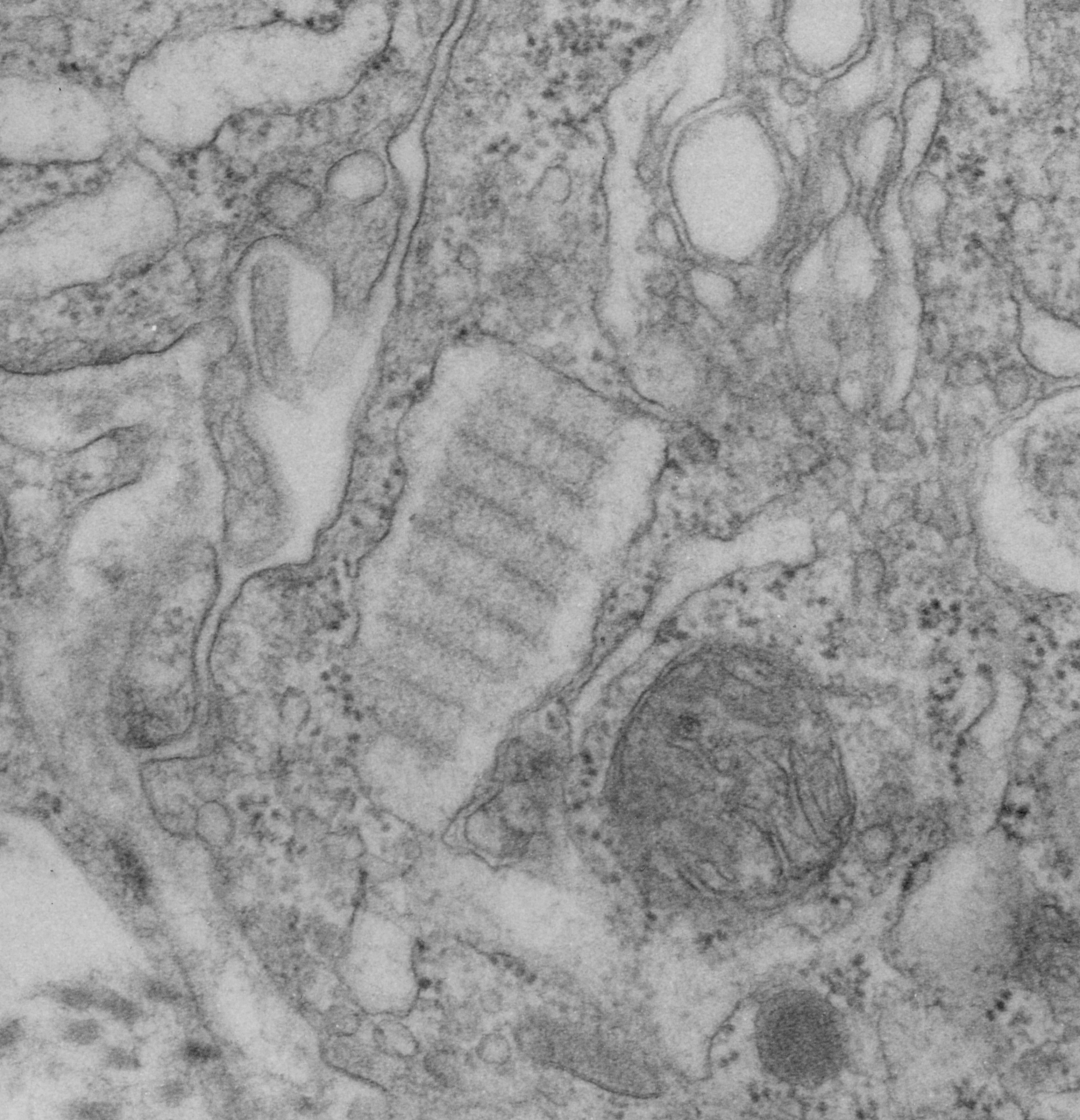
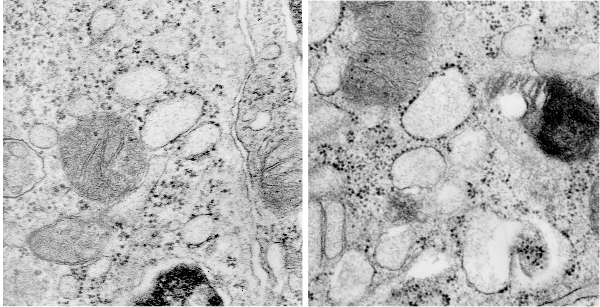
nb. since this blog, I found a similar micrograph of dog lung, in a paper by Stephens, Freeman, Stara and Coffin, Am J Path, 73: 711-726, 1973 which identified a very similar structure (cuboidal organization) in endothelial cells (they say)(Figs 14-16 in their publication). I cannot identify the cell in my micrograph as an endothelial cell, but can confirm that there are dense coated vesicles (at least 11 in this image) just like they found in their micrograph., and one edge of the cell in my micrograph is adjacent to collagen fibers. This would make the cell either epithelial or endothelial. It has no lamellar bodies and no nuclear profile, and only two kind of iff-ish mitochondria.
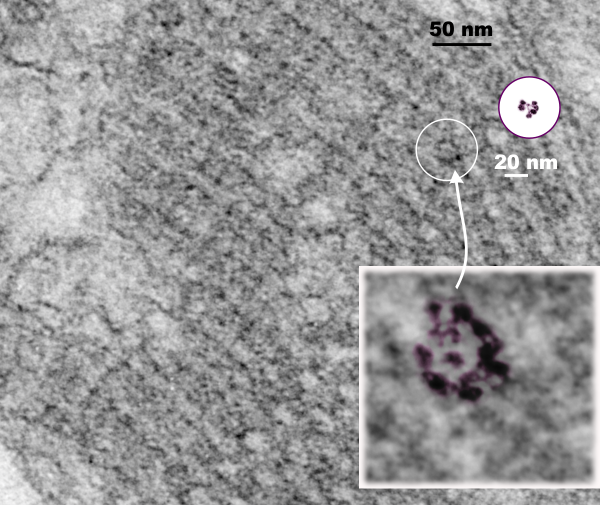
Circles identify the two molecules (one SP-A according to Voss et al, the other a similar sized protein (could be an octadecamer?? just making a guess here) or some other order of –merization. In the two images (see previous post) there were literally hundreds of times that one could visualize a bouquet with center point as the one enlarged in the inset, and pseudocolored a purple. One of the perplexing things is that when one begins to see structures in a repeated design like this…. many more can be pointed out, and then one wonders what is imagination and what is fact (that is the great tendency of the human brain to become immediately biased).
 So after finding this in the publication by Stephens et al, 1973, I decided to scan another area of that same cell and enlarge a portion, and highlight some structures (as above) which look like a multimer of SP-A in a lattice formation and post it here again. Would love to say I could count 18 blobs to depict the carbohydrate recognition domains (haha). I highlighted the “spokes” found in the inclusion below, I should have seen them in the inset inclusion above, but did not highlight them… however, they are there.. maybe 6 of them in top, maybe 7 in bottom inset. The configuration here , globby tops, skinny necks and central dot is pretty reminiscent of how the bouquet of SP-A is diagrammed.
So after finding this in the publication by Stephens et al, 1973, I decided to scan another area of that same cell and enlarge a portion, and highlight some structures (as above) which look like a multimer of SP-A in a lattice formation and post it here again. Would love to say I could count 18 blobs to depict the carbohydrate recognition domains (haha). I highlighted the “spokes” found in the inclusion below, I should have seen them in the inset inclusion above, but did not highlight them… however, they are there.. maybe 6 of them in top, maybe 7 in bottom inset. The configuration here , globby tops, skinny necks and central dot is pretty reminiscent of how the bouquet of SP-A is diagrammed.