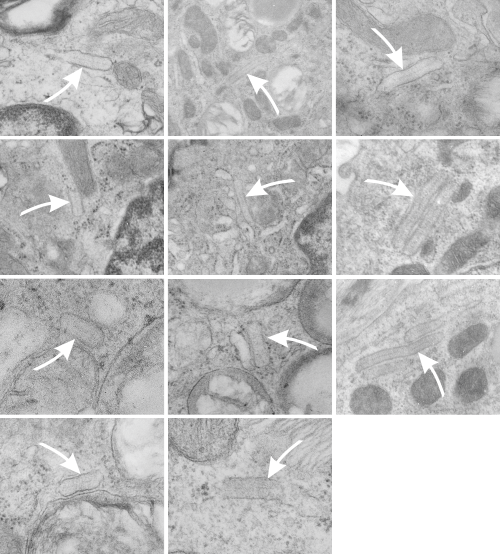
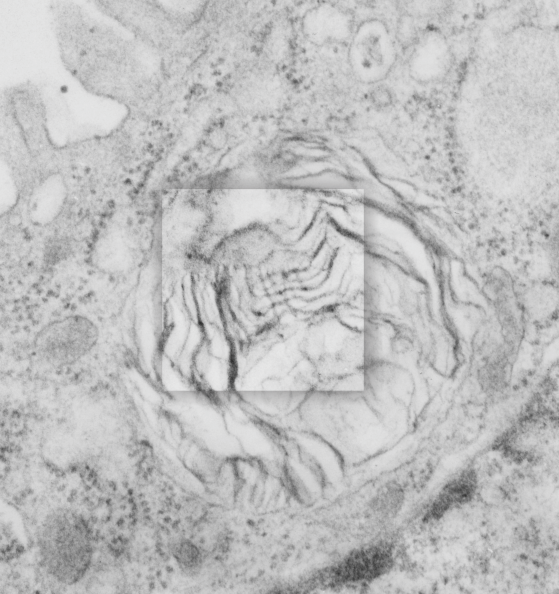
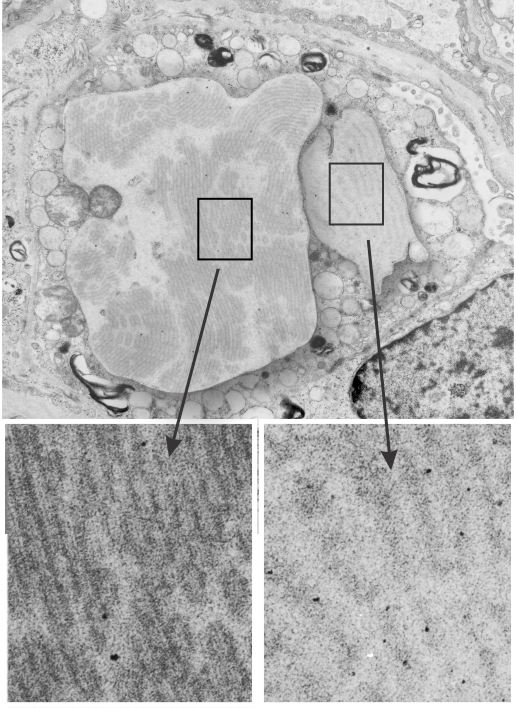
This post is something of a revelation, I think I knew this stuff from all the last 9 months of working on Surfactant protein A (SP-A) but it was pretty much exciting to think about all the times I have recognized its function while it was still unknown to me from a scientific standpoint.
Beginning with the observation (of about 30 years duration) that I rarely got upper respiratory infections. Why? you ask. It seemed to me to be directly related to my regular practicing the flute. I am not accomplished at music, but I did practice half hour or so, and at some points more, each day, and also the timing of my practicing might be important, as that was usually just before bedtime (specifically after the kids were in bed — or at least lying on the bed where I was positioned at the foot on the floor trying to multitask… stretching the leg tendons while playing Bach. ha ha. This resistance to upper respiratory tract infections was a real phenomenon in my mind, and I have understood it to be related to playing the flute for many years.
I also remember some “old wives” medicine tips…. that sounded like this. “If you are asthmatic, learn to play the clarinet” This was observational medicine, no question of its validity.
I stopped playing the flute a few years back, but took up running (upon the challenge from my daughter (39 years my junior) to run a marathon. Then we shaved the idea down to a half marathon (which we did both complete), and I kept running more or less 5 times a week. I still didn’t get upper respiratory infections very often… maybe a little more than before, but age is a variable too. So aerobic exercise (running) took the place of my “inhale exhale” exercise from the flute.
So also, taking up the study of strange layered intracisternal protein granules in the alveolar type II cells in the lung of a couple of mammalian species has taught me much about pulmonary biology. SP-A is an awesome protein, highly conserved, and really really important for innate immune responses… helping with so many of the “wellness” functions that occur in the lung, not just what its name would suggest, being a surfactant and lowering surfact tension in the alveolar spaces. So. here is the thing….PREVENTION is the best MEDICINE — go exercise yourself, or play the flute, or clarinet, or french horn, or tuba, or trumpet, or or or…but inhale “life” deeply, stretch out that chest, breath hard, exhale in a hurry, do something aerobic. In doing so, you force those type II cells to export those lamellar bodies which contain surfactant, which includes a significant amount of SP-A). The latter then helps opsonize all those viruses and bacteria, and other stuffs that cause disease, helps bring surfactant back into the type II cells for recycling, and helps move the junk into alveolar macrophages for “removal” from the lung, modulates inflammation and antigen processing…. your whole health will improve. All that SP-A is there for a reason, exercise your lungs, use that SP-A, be well. — and it is cheaper than the MD, and safer than the Scripts, your brain will thank you too, and it is there for you BUT ONLY IF YOU EXERCISE IT.