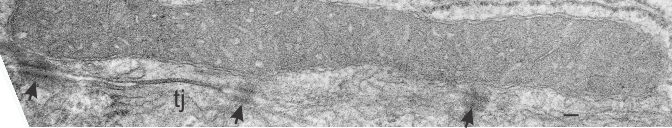
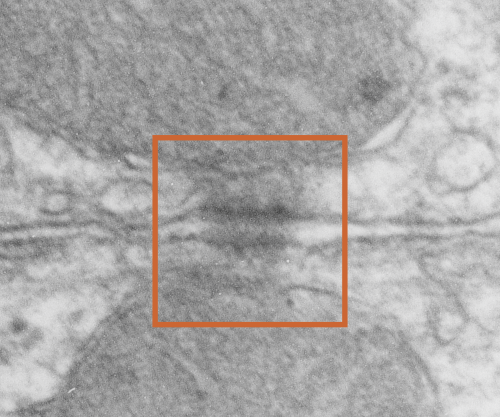
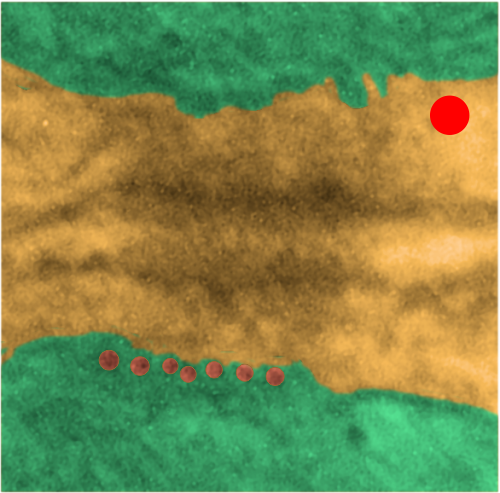
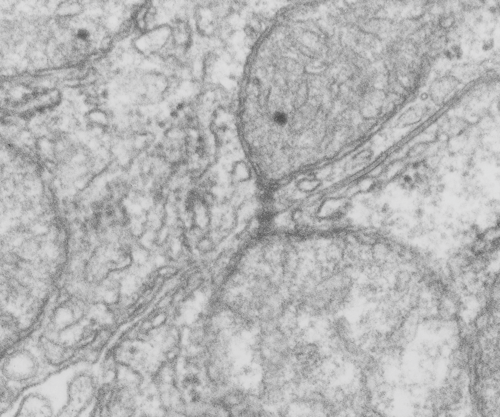
While perusing hundreds of electron micrographs of alveolar type II cells in half a dozen species i was disappointed NOT to see any desmosomes, or if I saw them they were not very well developed and tangentially cut, but I really think they may exist in a different form than they do, at least, in liver.
This I surmise from the fact that very little of the alveolar type II cell is actually “tethered” to other cells. A large portion (basal plasmalemma) abuts basement membrane, a tiny bit is close to alveolar type I cells, that connection by the way is very thin and would not even provide enough space wherein a mitochondria could reside and be found tethered to a desmosome, if i could find a desmosome. The large (perhaps the largest) plasmalemmal surface is in the alveolar space and no desmosomes possible there. So the hunt for desmosomal mitochondrial tethers in alveolar type II cells just came up empty. Empty for desmosomes, empty for desmosomal mitochondrial tethers, but of course there are plenty of mitochondria per se…. just not tethered to anything resembling a desmosome. I don’t even have an image of something which might be considered “close”.
It makes sense in terms of physiology however not to have rigid structures tightly binging alveolar type II cells together…. the tremendous movement within the alveolus durin inspiration and expiration would make it a little untenable to have to use energy, move mitochondria, make and break associations within the cell and the intermediate filaments, and lose the intercellular bonds in the cadherin molecules 20 or 30 times a minute. (and remember that is slow compared to the inspiration and expiration of some animals). So peripheral lung was probably a silly tissue to hunt for desmosomal mitochondrial tethers (there are other junctional complexes of course to prevent fluid from coming into the intercellular space that are easily seen, tight junctions and adherens junctions. Some describers of peripheral lung epi dont even mention desmosomes. HERE (i have never heard of this journal, so reader beware)