A modified rivet
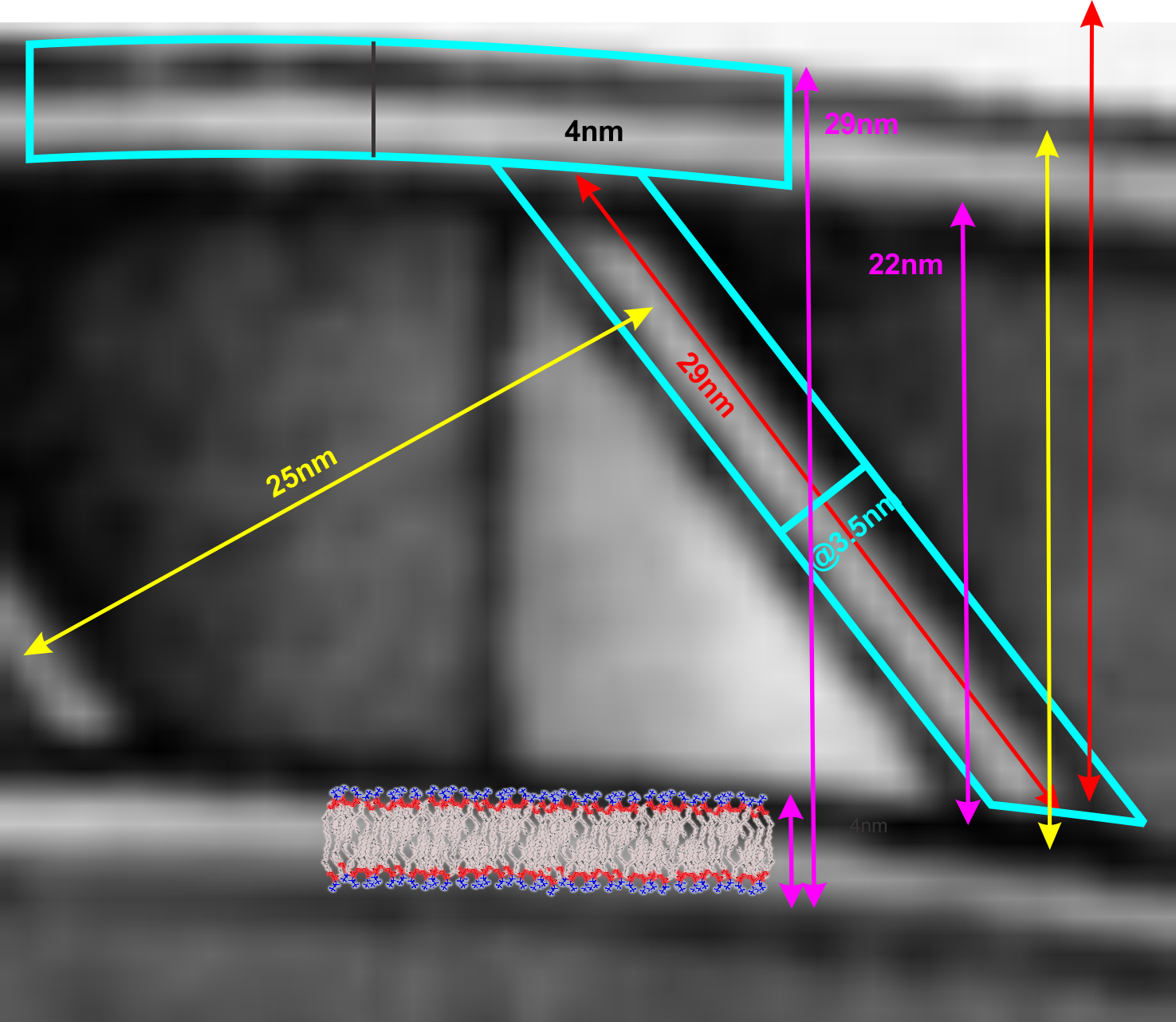
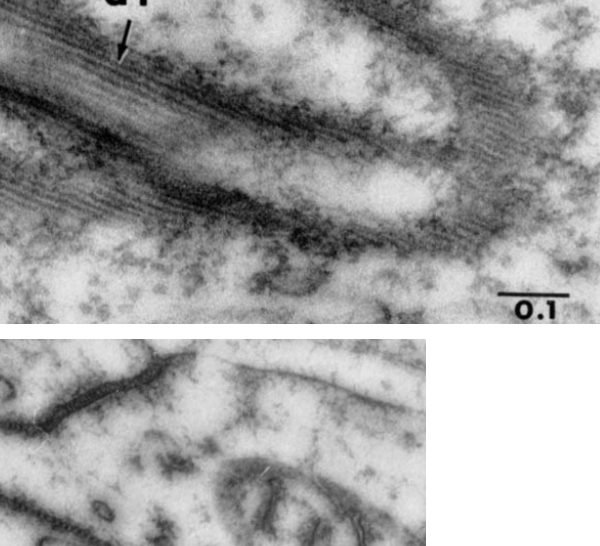
There are many names which have been given to these adhesion points, that is the desmosomes. They occur as spots, yes, of about 200-500nm in diameter, but they are certainly not spot”welds” which would imply that the two cell membranes involved become ONE, ie, the surfaces of the two sides being joined are changed. (This is more reminiscent of the tight junction, matching the spot-weld description somewhat but totally unlike a desmosome).
Others have called the desmosome a BOLT. This isn’t accurate either because the central line in the extracellular space of the desmosme is more like a velcro joint and not a solid inflexible mass but it is a moveable collection of various assemblies of cadherins – mainly desmocollins and desmogleins) where the center portion of the juncture (where the cadherins are bound in a central dense line with a distinct periodicity (something around 6-8nm) can be made and broken depending on ion concentration and cell signaling. This is not at all like a solid bolt-like structure, instead it is more like a zipper. Presumably the bolt analogy which referrers to a something “imagined” but not seen with electron microscopy was just a careless comment since a desmosome penetrating both sides of two adjacent cells is certainly NOT SOLID. A bolt is not only solid but is temporary (or can be temporary) this on the other hand is more like a desmosome.
The desmosome is made up of hundreds of molecules positioned parallel and perpendicular to the plasmamembrane, allowing for flexibility and resistance to various stresses: shear, cleavage, tensile and peel, compressive and torsion. I am sure there are others. The desmosome might be a partial solution to shear, but it would be single shear, since there are only two surfaces involved, one from each adjacent cell.

and ensuring the adhesive molecules penetrate the proximate side of the adjacent plasmalemma of both nearby cells. The bolt analogy doesn’t work since the central joint of the desmosome is easily “made” and “broken” depending upon cell signaling, stage of cell cycle, calcium concentration and sensibility of the cadherin molecules, ie. whether the desmosome is going to be removed or substituted, or unzipped.
- semi-permanent
- cylindrical shank with “head” within each of the two adjacent cells
- similar to a buck at the intercellular junction between the cadherins from each of the two cells.
- major differences is that the “buck” is two sided, like the velcro…. and the integrity of the attachment sis maintained by calcium concentration (and likely many other as yet unknown factors).
A rivet however is defined as a “permanent fastener” so here we have a problem right away…. desmosomes are made and unmade on a regular basis, moved from here and reassembled there to accomodate changes in, activity, stage of the cell cycle, size, forces, maturity of the cells, and countless others. In fact all junctions have to be totally responsive to intra and extracellular events.
Is a desmosome a little like a pop nut, or hollow wall hanger, where the is flexibility in the tethering of cadherins…. just a little, and also in tethering the intermediate filaments by desmoplakin?
Or is the desmosome is more like a bilateral (two headed) unzippable ductile lap rivet.
One of the unique features of attaching two cells with a desmosome that has no parallel in bringing two pieces together in a rivet, is that both sides in the rivet process need to be accessed….which in building….can present a problem. However, intracellular activity makes this access routine and the intercellular components are managed from the inside of each of the respective cells.
I have thought for a while about the arrangement of molecules, and its very nice parallel+perpendicular+parallel+perpendicular line up and felt that this was certainly better than any man-made adhesion rivets…. I wondered whether such a complex set of parts could actually be developed into a real live-functional construction friendly rivet.
Brazing or soldering joint….not applicable in my opinion.
Types of joints….. the desmosome is NOT a butt joint, clearly a lap joint. The spacing of desmosomes just like rivets, will be dependent upon many biological parameters, including states of differentiation.