Some measurements, just made for my own information, might be useful in a relative sense to others looking at nucleus, nuclear architecture, and nucleolar architecture. Some data exist of course, but In the dataset that I am putting together from many different experimental protocols (archived) might lead to some new insights on numerical density of nuclear pores during times of cell stress, apoptosis, and in previously studies KO mice. The numbers are meant to be “observations” and a suggestion.
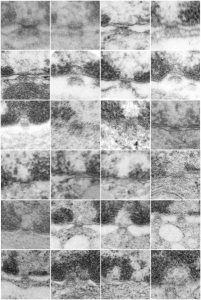
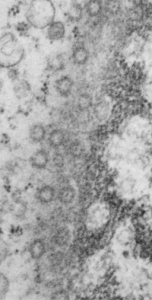
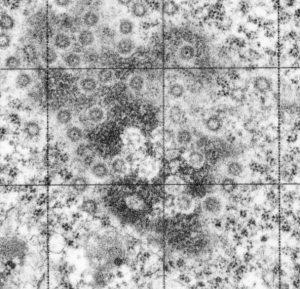
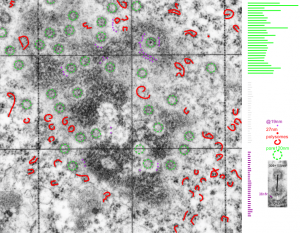
This particular diagram has as its background (top image) unretouched, and from a group of micrographs which were used for morphometry (hence the ink jet grid on the surface). It is hepatocyte, from a +/- 14CoS mouse which did receive NTBC (as a control for experimental mice receiving NTBC (data here) but showed pretty normal histology – but I predict with enough time and scrutiny something would show up. Octameric nuclear pores are shown as green dotted circles, ribosomes and polysomes presumably attached to the outer nuclear membrane, red, ribosomes, red = @27 nm diameter, peri-pore chromatin beads diameter, @19-20nm. Condensed chromatin at the inner nuclear membrane is well defined and “wound” in many places as a fibril also, possibly around 20+ nm diameter but longer lengths. About %66 of nuclear pores in this photo have central densities indicating some molecule is in transport one way or the other.
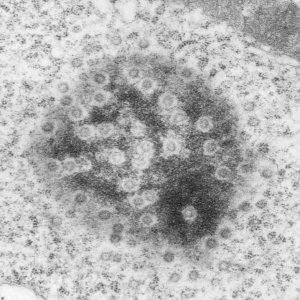
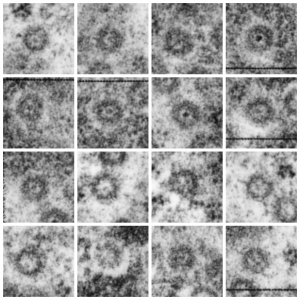
This particular animal had a great tangential section showing a large portion of the nucleus studded with nuclear pores and I used that opportunity to make some measurements on the diameter of the chromatin with beads (@19nm)-on-a-string appearance adjacent to the nuclear pores but outside the chromatin exclusion area around the pores and the distance between those beads (38nm); to measure the width of the chromatin exclusion area at places around the nuclear pores (maybe 2 -5 measurements perpendicular between pore and chromatin, per pore) (60nm+ 7nm and to measure the pore to pore distances as well (n=27, MEAN, 309nm+ 109nm (SD)).
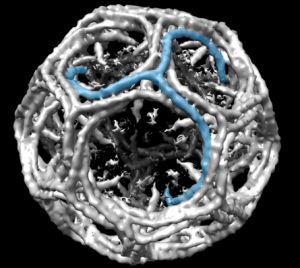
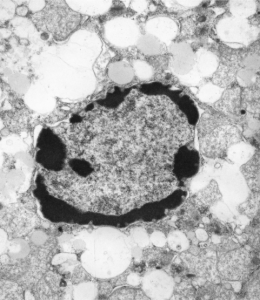
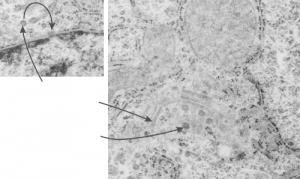 Might be a section of a clathrin coated vesicle…. when i google it, this wonderful model came up. Sure looks (at 100nm) like it could work. What a marvelous amount of research has provided the molecular structure (triskelion) shown by them (on wikipedia) in blue.
Might be a section of a clathrin coated vesicle…. when i google it, this wonderful model came up. Sure looks (at 100nm) like it could work. What a marvelous amount of research has provided the molecular structure (triskelion) shown by them (on wikipedia) in blue.