There is one guinea pig in this group of animals that was completely different, in the sense that all the surfactant granules within alveolar type II cells in this animal are just overpowering all other cytoplasmic elements. This was a control animal which makes for interesting speculation, maybe even an oblique reference to the fact that this structure is a response to environmental influence (in this case probably a massive pulmonary infection or immune disorder. At least there was no experimental reason for alveolar type II cells to display so many granules.
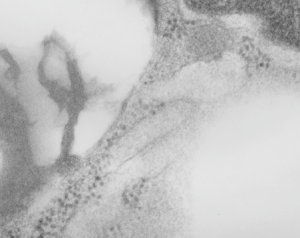
Posted previously are images of an interesting variation on the granules, those which, in between the 100 nm layering pattern that is most prominent, some fuzzy rounded ball-like groups of protein exist. A previous post that mentions these structures, in that case they were given a dimension of 50-60 nm… here I measured them more like 49 nm… so basically the same. I was wondering at that time if they were surfactant protein A or not. I am wondering now if they might be surfactant protein D. Well, still leaning toward SP-A, in a configuration that would be two molecules “deep” or half the width of the banding seen in the granules.
I did find this reference from Crouch et al, 1994 which had rat SP-D images that were awesome, however the dimensions don’t quite fit… and they give 46 nm as the size for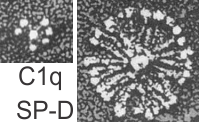 a single arm of the SP-D molecule, and that is exactly half of what I measure as the full diameter of one of these dense structures diameter (47 nm). They calibrated their microscope well, I am not arguing with their final dimensions. From their paper are two screen shots C1q (left) and SP-D (right). The magnification is not given in a micron marker bar, but C1q is just a little smaller than SP-A, there fore if the relative magnifications are the same in these two, then that makes SP-D about twice as big as SP-A would be so maybe 50 microns which sort of contradicts the size given. If C1q has a spread of the CRD close to SP-A (which is 20-20 nm) and picture on left is 20-30 nm diameter, then it appears that the SP-D here is about 2.5 times larger than SP-A would be and that is more like 70 nm or even 100 nm, and a previous post on this blog calculated the round SP-D at 100
a single arm of the SP-D molecule, and that is exactly half of what I measure as the full diameter of one of these dense structures diameter (47 nm). They calibrated their microscope well, I am not arguing with their final dimensions. From their paper are two screen shots C1q (left) and SP-D (right). The magnification is not given in a micron marker bar, but C1q is just a little smaller than SP-A, there fore if the relative magnifications are the same in these two, then that makes SP-D about twice as big as SP-A would be so maybe 50 microns which sort of contradicts the size given. If C1q has a spread of the CRD close to SP-A (which is 20-20 nm) and picture on left is 20-30 nm diameter, then it appears that the SP-D here is about 2.5 times larger than SP-A would be and that is more like 70 nm or even 100 nm, and a previous post on this blog calculated the round SP-D at 100  nm. I am opting for the fuzzy balls in the granules of alveolar type II cells in this study to be SP-A. Per diagram.
nm. I am opting for the fuzzy balls in the granules of alveolar type II cells in this study to be SP-A. Per diagram.
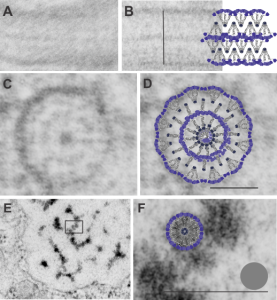
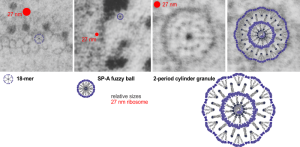
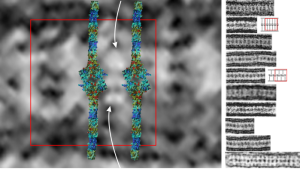
The image below (9124_17082_gpig_301_alveolar type II cell with granules, lamellar body (upper left under the legend) and a fuzzy all inclusion (in the RER i believe). I saw these fuzzy balls embedded within granules but this is the only time I saw a cluster of them as a separate granule. Image below show the fuzzy balls clustered on the right, the figure below that shows relative sizes of ribosomes (red dots of 27 nm) and some examples of the diameter of the fuzzy balls at something around 47 nm and an optimally sectioned fuzzy ball (greatest diameter) of almost 50 nm.
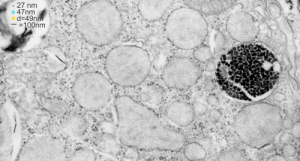
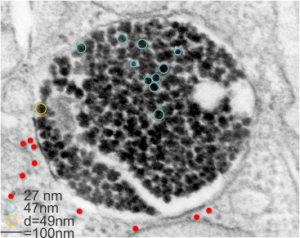
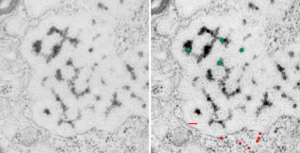
These fuzzy balls can occur, per above comments, within the granules which are otherwise typically without these spherical densities. micrograph 9864_17085_gpig_301. The red dots on right are more or less equivalent to 27 nm diameter (ribosome size) and the red bar is therefore equivalent to 100 nm. Because of the lack of prominent layering and the added width (red bar) for the periods, it is presumed that this represents a tangential cut, and the size of the electron dense round balls (blue circles in images on right) is something around 47 nm (which is similar to estimates in other micrographs based on the size of a ribosome. So these fuzzy balls can be seen both as separate membrane bound entities or found between the period layers of granules. Unretouched photo on the left, photo on the right has the end-on single circular profiles highlighted…. looking like bulls-eyes.